Description
CD30L: A crucial co-stimulatory driver that amplifies T-cell activation
- Gene Information: TNF Superfamily Member 8 (TNFSF8, also known as CD30L) is a protein-coding gene located on chromosome 9q32-q33.1. This cytokine is a ligand for receptor TNFRSF8 (also known as CD30).
- Protein Expression: CD30L is rarely expressed under resting conditions and is predominantly localized on the surface of stimulated and activated immune cells. Activated T cells serve as the primary source of CD30L expression, though it is also expressed on activated B cells, monocytes, macrophages, and dendritic cells.
- Signaling Pathway: The binding of CD30 and CD30L forms a trimer, which can generate signal transduction for binding with TRAF members, induce the activation of NF-κB and MAPK pathways, and participate in the processes of tumor cell proliferation and apoptosis, as well as the activation and proliferation of T and B cells.
- Therapeutic Inhibition: Blocking the interaction between CD30 and CD30L can abrogate the co-stimulatory signals driving immune cell hyperactivation, thereby ameliorating autoimmune inflammatory responses.
Targeting strategy
CD30L
- The exons 1-4 of mouse Cd30l gene that encode extracellular domain were replaced by human counterparts in B-hCD30L mice.
- The genomic region of mouse Cd30l gene that encodes transmembrane domain and cytoplasmic portion was retained. The promoter and 5’UTR region of the mouse gene were also retained. The 3’UTR region of the mouse gene are replaced by human counterparts. The CD30L expression was driven by endogenous mouse Cd30l promoter, while mouse Cd30l gene transcription and translation will be disrupted.
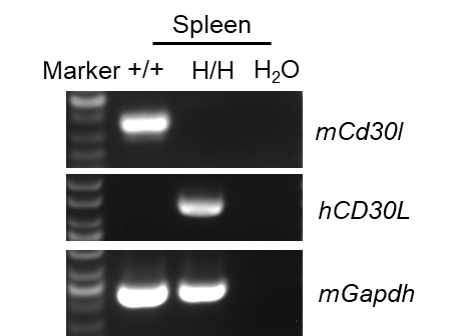
CD30L mRNA Expression Analysis in Spleen
- Human CD30L mRNA was specifically and correctly expressed in B-hCD30L mice.
Species specific analysis of CD30L gene expression in wild-type C57BL/6JNifdc mice and homozygous humanized B-hCD30L mice by RT-PCR. Spleen was collected from wild-type C57BL/6JNifdc mice (+/+) and homozygous B-hCD30L mice (H/H). Mouse Cd30l mRNA was only detectable in wild-type C57BL/6JNifdc mice. Human CD30L mRNA was only detectable only in homozygous B-hCD30L mice, but not in wild-type C57BL/6JNidfc mice.
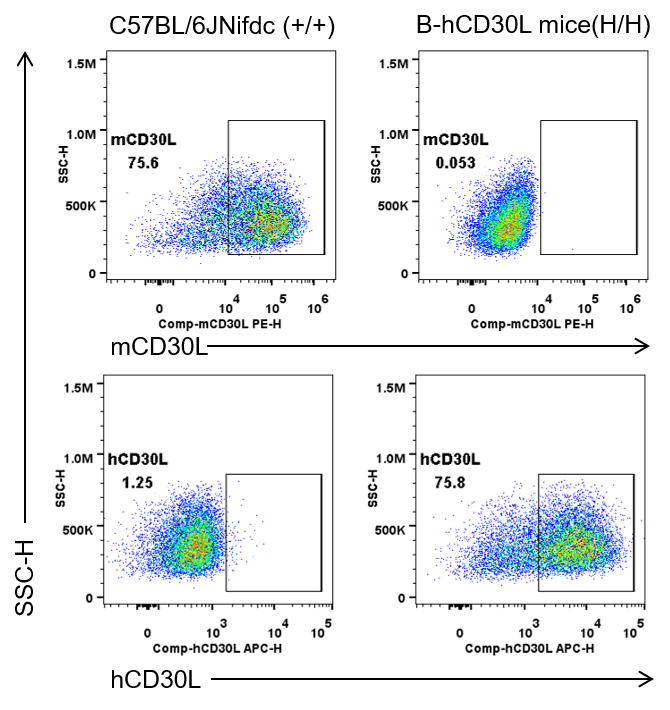
CD30L Protein Expression Analysis in Spleen
- Mouse CD30L was exclusively detectable in T cells of wild-type C57BL/6JNifdc mice. Human CD30L was exclusively detectable in T cells of homozygous B-hCD30L mice, but not in wild-type C57BL/6JNifdc mice.
Strain specific CD30L expression analysis in wild-type C57BL/6JNifdc mice and homozygous humanized B-hCD30L mice by flow cytometry. Splenocytes were collected from wild-type C57BL/6JNifdc mice (+/+) and homozygous B-hCD30L mice (H/H). Splenocytes are incubated in a medium containing Cell Activation Cocktail (without Brefeldin A) (Biolegend, 423303) before analysis of CD30L surface expression on gated T cells. Protein expression was analyzed with anti-mouse CD30L antibody (Biolegend, 106405) and anti-human CD30L antibody (RD, FAB1028A) by flow cytometry.
Functional Validation
- PRA052 analog (in house) could reduce the percentage of IL17A+ CD4+ T cells and decrease the production of mouse IL17A and mouse IL23 in B-hCD30L mice during Th17 cell differentiation.
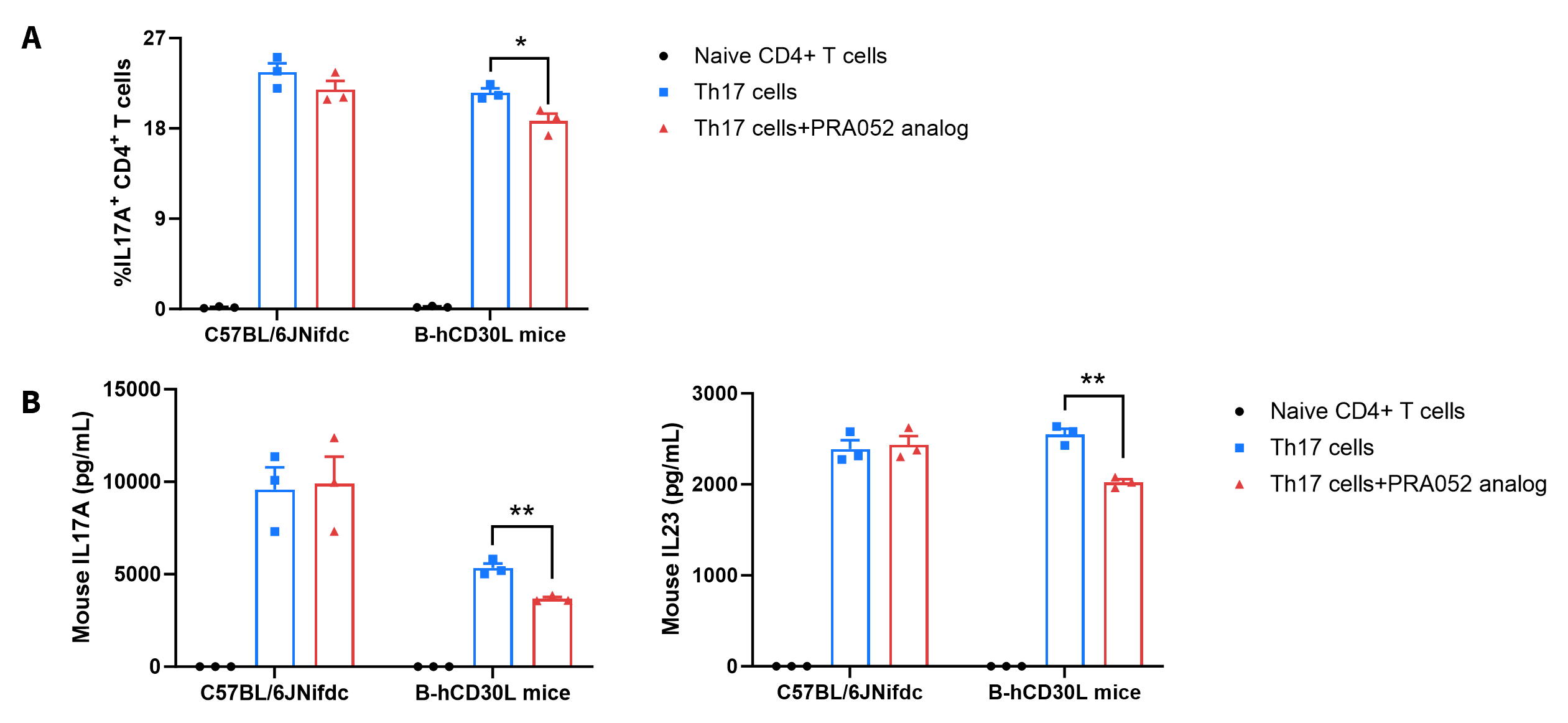
Ex vivo functional analysis in wide-type C57BL/6JNifdc and homozygous B-hCD30L mice. Naïve CD4+ T cells were sorted from the splenocytes of wild-type C57BL/6JNifdc mice (+/+) and homozygous B-hCD30L mice (H/H), and were assessed after 72 h of incubation with rmIL6, rmIL23, rmTGF-β1, anti-mouse IFN-γ antibody and anti-mouse IL4 antibody in combination with bead-associated CD3 and CD28 mAbs. (A) The percentage of mIL17A positive cells were assessed by flow cytometry. The anti-human CD30L antibody PRA052 analog (in house, 50 μg/mL) could reduce the percentage of IL17A+ CD4+ T cells in B-hCD30L mice during Th17 cells differentiation, as PRA052 analog only recognizes human and does not recognize mice. (B) The protein expression of mouse IL17A and mouse IL23 in cell culture supernatant were measured by ELISA. The anti-human CD30L antibody PRA052 analog (in house, 50 μg/mL) could reduce the production of mouse IL17A and mouse IL23 in B-hCD30L mice during Th17 cells differentiation. Values are expressed as mean ± SEM. Significance was determined by unpaired t test. *P < 0.05, **P < 0.01.
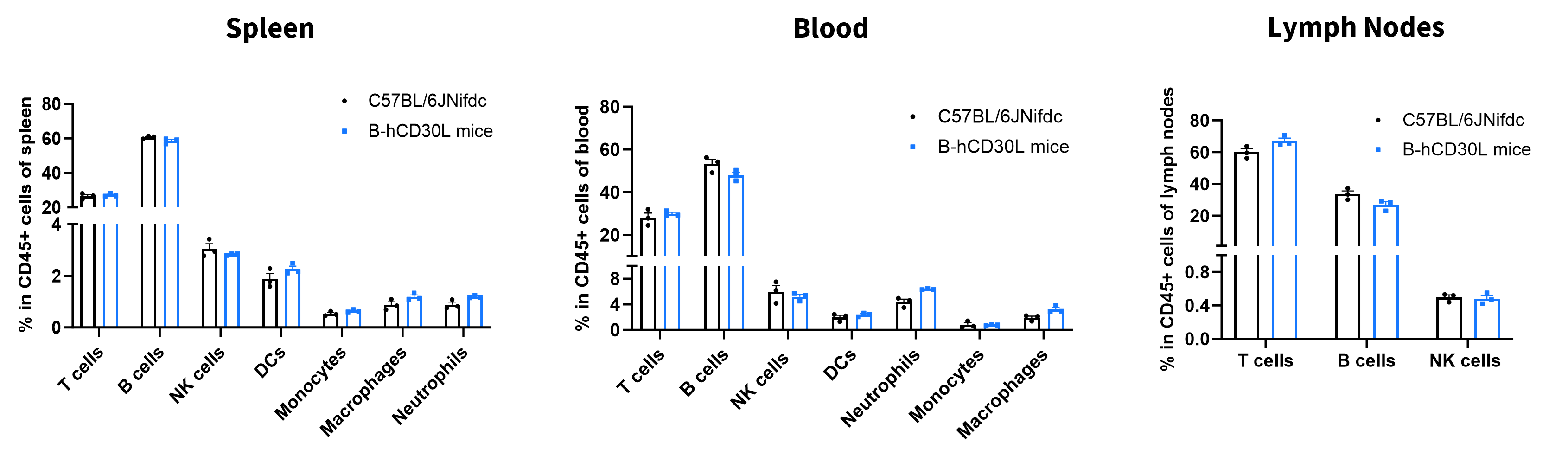
Analysis of Leukocyte Subpopulations
- The percentages of T cells, B cells, NK cells, DCs, monocytes, macrophages, and neutrophils in homozygous B-hCD30L mice were similar to those in C57BL/6JNifdc mice.
- Humanization of CD30L does not affect normal immune cell development or splenic distribution.
Analysis of leukocyte subpopulations by flow cytometry in immune organs and blood. Splenocytes, peripheral blood, and lymph nodes were isolated from female C57BL/6JNifdc and B-hCD30L mice (female, 6-week-old, n=3). Single live cells were gated on the CD45⁺ population and analyzed by flow cytometry as indicated. Values are expressed as mean ± SEM.
Analysis of T Cell Subpopulations
- The proportions of CD4⁺ T cells, CD8⁺ T cells, and Tregs in homozygous B-hCD30L mice were comparable to those in C57BL/6JNifdc mice.
- Humanization of CD30L does not affect normal T cell development, differentiation, or splenic distribution.
Analysis of T-cell subpopulations by flow cytometry in immune organs and blood. Splenocytes, peripheral blood, and lymph nodes were isolated from female C57BL/6JNifdc and B-hCD30L mice (female, 6-week-old, n = 3). Single live cells were gated on the CD3⁺ T-cell population and analyzed by flow cytometry as indicated. Values are expressed as mean ± SEM.
In Vivo Efficacy of Anti–hCD30L Antibody in T-Cell-Dependent Antibody Response (TDAR) Assay
- PRA052 analog (in house) could reduce the percentage of IL17A+ CD4+ T cells and decrease the production of mouse IL17A and mouse IL23 in B-hCD30L mice during Th17 cell differentiation.
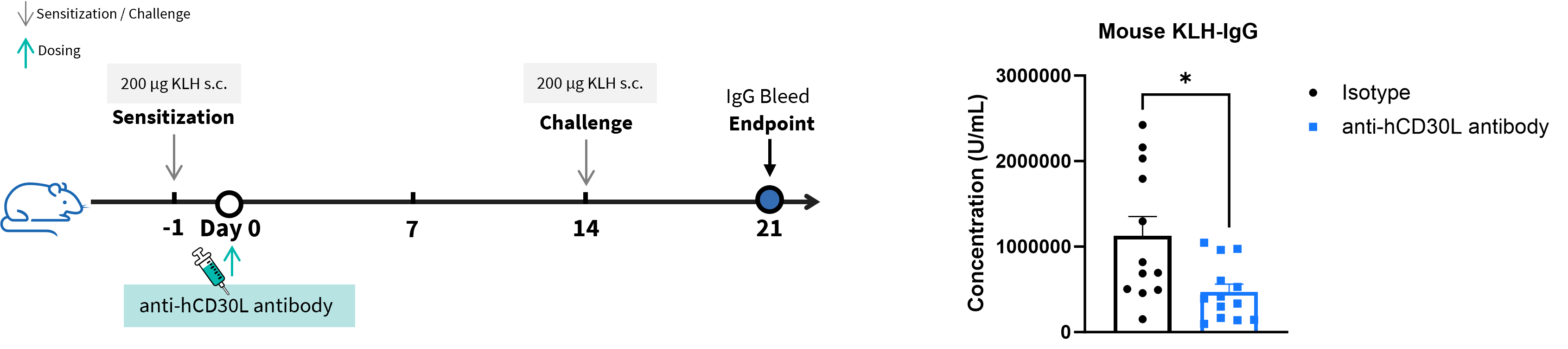
T-cell-dependent antibody response assay in homozygous B-hCD30L mice. B-hCD30L mice were used for the TDAR assay to evaluate the efficacy of the anti-hCD30L antibody (provided by the client). On Day 0, the mice were subcutaneously injected with 200 μg of KLH for immunization, another subcutaneous injection of 200 μg KLH was administered for boosting on Day 14. On Day 21, serum was collected from the mice to assess the levels of KLH-specific IgG. Treatment with the anti-hCD30L antibody (provided by the client) significantly reduced the level of KLH-specific IgG in mouse serum, indicating that the anti-hCD30L antibody could block T cell-dependent B cell activation. This data validated the effectiveness of B-hCD30L mice as a preclinical evaluation model for anti-hCD30L antibodies. Values are expressed as mean ± SEM. Significance was determined by unpaired t test. *P < 0.05.
Note: This experiment was conducted by the client using B-hCD30L mice.
* When publishing results obtained using this animal model, please acknowledge the source as follows: The animal model [B-hCD30L mice] (Cat# 113224) was purchased from Biocytogen.